RNA干扰技术原理、机制及应用
RNA 干扰是由 RNA 分子介导的基因沉默调控机制。该机制通过 siRNA、miRNA 等非编码 RNA 与靶基因 mRNA 发生序列特异性结合,进而介导靶 mRNA 降解或抑制其翻译过程,最终实现靶向基因表达沉默与功能抑制。
RNA干扰技术通常涉及到以下几种RNA:
①miRNA(micro RNA),小分子核糖核酸/微核糖核酸,真核中广泛存在的一类长约22nt(21-24nt)的有茎环结构的RNA分子,成熟miRNA为单链,可诱导标基因的mRNA降解或抑制其蛋白质翻译来调控目的基因的表达。

miRNA结构示意图
②siRNA(small interfering RNA),小干扰核糖核酸,是一类长约19-25nt(实验中通常为19nt或21nt,干扰效率较高)的双链RNA分子,一般3'端有两个游离碱基,可使mRNA降解。
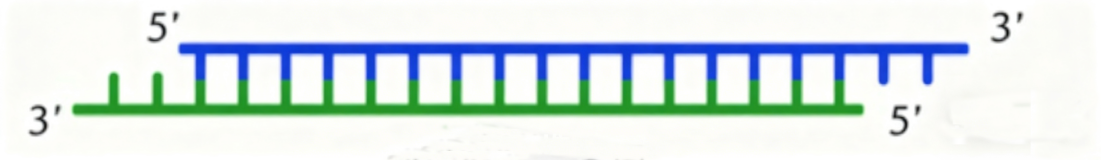
siRNA结构示意图
③shRNA(short hairpin RNA),小/短发夹RNA,依赖茎环结构产生的短双链RNA,长约19-25nt,在细胞内经酶切产生siRNA。
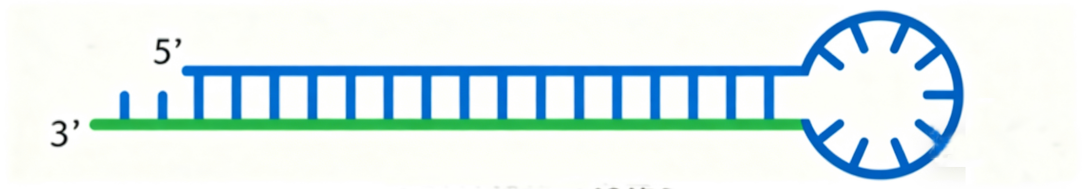
shRNA结构示意图
最终发挥沉默效应的主要是siRNA和miRNA
siRNA和miRNA干扰机制
miRNA翻译抑制或基因沉默机制:
miRNA由内源性基因转录而来,其初级转录本(pri-miRNA)在细胞核中被Drosha和DGCR8处理形成前体miRNA(pre-miRNA)。随后被Exportin 5转运到细胞质后,pre-miRNA被Dicer酶进一步处理成为成熟miRNA。
成熟miRNA链与Ago1-Ago4蛋白结合形成RNA诱导沉默复合体(RISC)。
当 miRNA-RISC 复合物与靶 mRNA 的 3'UTR 发生不完全互补配对时,主要通过阻断核糖体结合、抑制翻译延伸等方式抑制蛋白质合成;若为完全互补配对,则可触发 Ago 蛋白的切割活性,导致靶 mRNA 降解,实现转录后基因沉默。
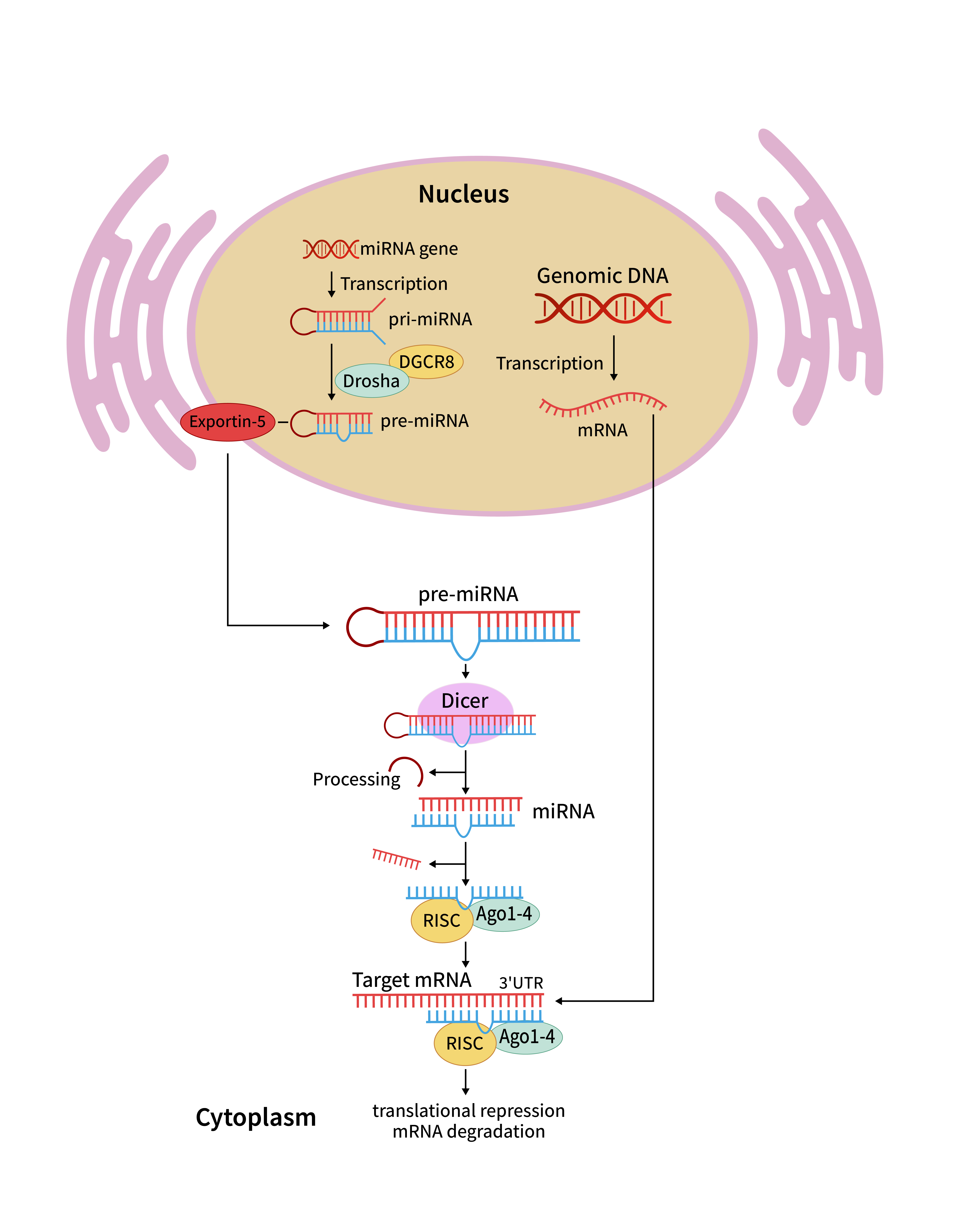
miRNA翻译抑制机理图
siRNA基因沉默机制:
合成的外源 siRNA 通过转染胞吞进入细胞,少量 siRNA 经溶酶体逃逸进入细胞质。siRNA 与 Dicer-TRBP 组成的 RISC 装载复合体结合,随后被转运至 Ago2 蛋白处,完成 RISC 复合物组装。组装后的 RISC 促使双链 siRNA 解旋,无功能的过客链随即降解,仅保留引导链。引导链与靶 mRNA 发生特异性互补结合,Ago2 利用内切酶活性切割靶 mRNA,断裂的核酸片段被胞内外切酶逐步降解,以此在转录后层面抑制基因表达,实现靶基因沉默效应。
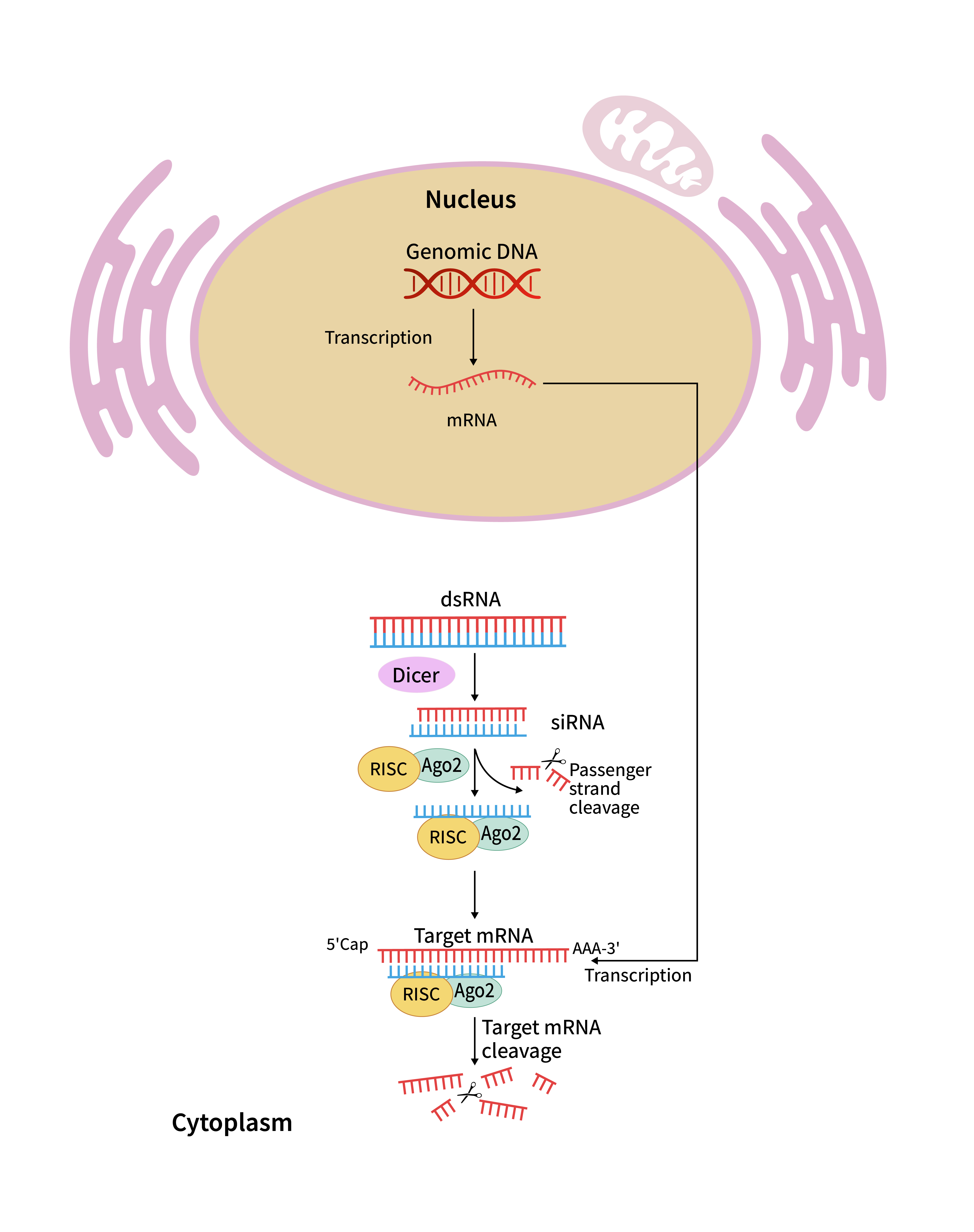
siRNA翻译抑制机理图
shRNA、siRNA和miRNA干扰工具的对比
| siRNA(小干扰 RNA) | shRNA(短发夹 RNA) | miRNA(微小 RNA) | |
|---|---|---|---|
| 来源 | 人工合成或外源导入 | 人工设计、载体转录 | 内源性、基因组编码 |
| 结构 | 双链(19–25 nt),3’端 2 nt 突出 | 单链发夹结构(含环 + 双链臂) | 单链(21–24 nt),不完全互补 |
| 互补性 | 与靶 mRNA完全互补 | 加工后为 siRNA,完全互补 | 与靶 mRNA不完全互补(3’UTR) |
| 作用机制 | 入 RISC→切割靶 mRNA→降解 | 核内转录→Dicer 切为 siRNA→同 siRNA 通路 | 入 RISC→抑制翻译或促 mRNA 降解 |
| 作用靶标 | 一对一(单一靶基因) | 一对一(单一靶基因) | 一对多(多靶基因,调控通路) |
| 作用时效 | 瞬时(3–7 天) | 长期稳定(可整合基因组) | 内源性、持续调控 |
| 主要应用 | 瞬时基因沉默、药物研发 | 稳定细胞系 / 动物模型、长期沉默 | 基因调控研究、疾病机制 |
实验选择建议
| 实验需求 | 推荐选择 | 原因 |
|---|---|---|
| 短期基因功能研究 | siRNA | 作用快速,适合瞬时转染实验 |
| 长期稳定沉默 | shRNA | 可整合基因组,稳定表达,适合构建稳定细胞系 |
| 难转染细胞 | shRNA | 病毒转导效率更高 |
| 研究内源性miRNA功能 | miRNA mimic/inhibitor | 模拟或抑制内源miRNA |
| 研究miRNA调控网络 | miRNA | 内源性持续调控,具有组织特异性 |
RNA干扰技术的应用
基础医学与生命科学研究;
医药治疗领域;
农业领域:绿色防控与品质改良。
 | Flora Flora是EnkiLife的技术支持专家,熟悉免疫学和神经科学,致力于为客户提供高品质的产品搭配和技术支持,以帮助实现神经退行性疾病和其它神经科学方面的研究。 |